
It is certainly fitting that in the year we celebrate the 200th anniversary of Charles Darwin’s birth and the 150th anniversary of the publication of his monumental work On the Origin of Species, a major advance in our understanding of human evolution should be announced.
After 15 years of painstaking study by 47 researchers, the journal Science has devoted its October 2 issue to reports, 11 papers in all, on the fossilized remains of what is interpreted to be an early form of hominin, the group including humans and all human ancestors back to the evolutionary split with the last common ancestor with chimpanzees.
The fossils, dating to 4.4 million years ago (mya), were discovered in the Afar Rift region of Ethiopia by a research team lead by Professor Tim White of the University of California at Berkeley. They have been classified under the taxonomic designation Ardipithecus ramidus. The initial discovery was made in 1992, consisting of several teeth and a jaw. A report of these finds was published in the journal Nature in 1994. Over the next year, additional discoveries were made, ultimately totaling more than 100 fossilized bone specimens, many quite fragmentary. Together, these represent a minimum of 36 individuals.
The most complete individual, an adult female, has been nicknamed “Ardi.” She is estimated to have been 120 centimeters (4 feet) tall and to have weighed 50 kilograms (110 pounds). Ardi is represented by a skull and teeth, as well as the pelvis, hands, and feet. She takes her place among the pantheon of fossil hominin individuals, including Lucy (Australopithecus afarensis), the Taung child (Australopithecus africanus), and the Nariokatome boy (Homo erectus), that illustrate important steps in hominin evolution.
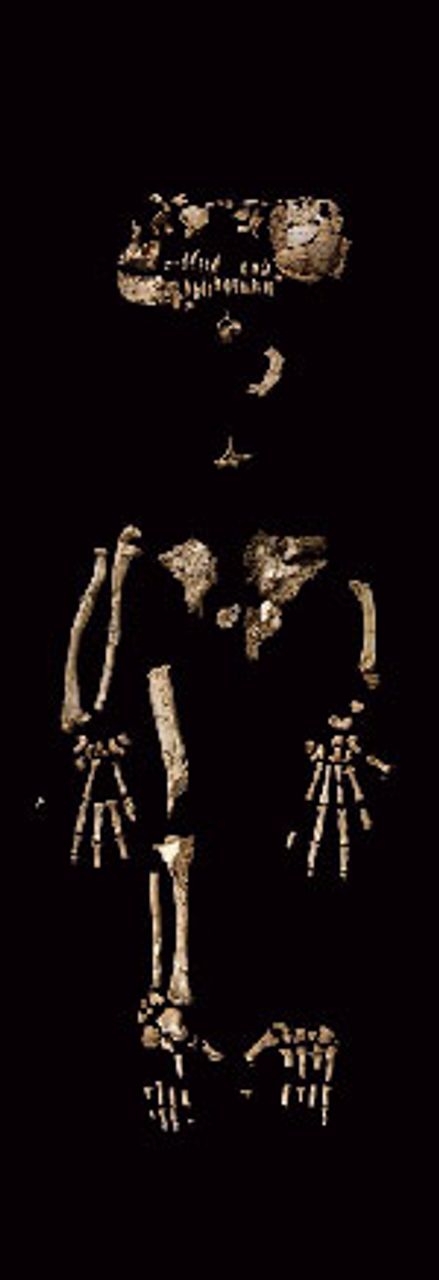 Figure 1. “Ardi.” All images
Figure 1. “Ardi.” All images are from Science
Until now, our knowledge of the hominin fossil record had been extremely meager before about 3.7 (mya), the age of the earliest known australopithecine remains. Analysis of DNA has indicated that modern humans and modern chimpanzees are closely linked in evolutionary history, and share approximately 98 percent of their genetic material. Using “molecular clock” rates of genetic change standard among primates and other mammals, scientists have calculated that the chimpanzee and human lineages diverged as early as 6 mya.
Over the past decade, new fossil evidence has moved this divergence back to 7 million years or more, following the discovery of what are purported to be very early hominins, Orrorin tugenensis and Sahelanthropus tchadensis. However, because these remains are fragmentary, their status is open to debate.
Knowledge of human evolutionary history prior to australopithecines has, therefore, remained extremely limited; indeed, half of hominin evolutionary history was largely unknown. This lack of evidence was exacerbated by the absence of a known fossil record for chimpanzee and gorilla ancestors. DNA data demonstrates that gorillas are slightly more genetically distant from humans than are chimps. The gorilla lineage split from the common hominin and chimp lineage before these latter two separated somewhat later.
Anthropologists have tended to envision very early hominins as rather “ape-like,” resembling modern chimps and gorillas. “Hominoids,” or what we commonly call the apes, are distinguished from monkeys by a suite of anatomical features well designed for a special kind of arboreal locomotion.
“Suspensory climbing,” which is practiced with an upright body posture and the grasping of multiple branches by multiple limbs, allows apes to grow larger in size while still remaining agile high up in the trees. This kind of climbing is present in all living apes except humans. Gibbons, siamangs, orangutans, gorillas and chimpanzees have arms longer than their legs; their wide torsos facilitate extended reach of limbs, and their short lower backs stabilize their upright torsos. Chimpanzees and gorillas use a special form of locomotion on the ground called “knuckle-walking.” Humans, by contrast, walk upright.
Some anatomical features of the torso and arms in modern humans attest to our ape-like heritage. But because we walk upright on the ground and no longer climb, we have lost some of the specializations that characterize other apes. However, elucidation of the precise sequence of evolutionary forms between arboreal and terrestrial adaptations awaited additional fossil discoveries.
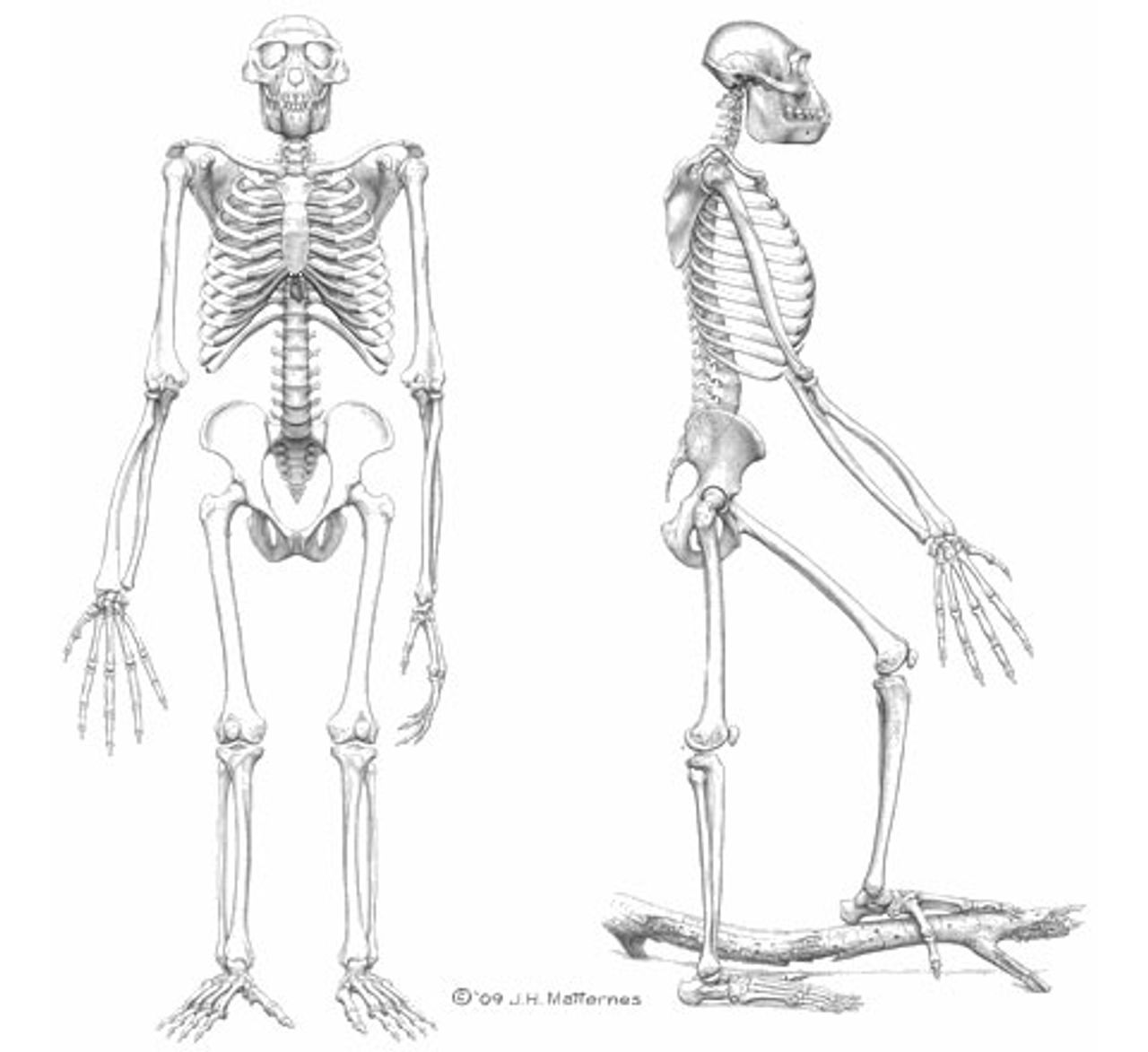 Figure 2. Artist’s rendition of a complete Ardipithecus skeleton
Figure 2. Artist’s rendition of a complete Ardipithecus skeletonThe recent publication of extensive research on the fossil remains of Ardipithecus ramidus helps fill the gap in the fossil record, and may provide the basis for a quantum leap in our understanding of early hominin evolution. In addition to describing the fossil remains of Ardipithecus ramidus, the authors of the articles in Science argue that Ardipithecus will revolutionize our understanding of the evolution of the apes.
By affording a view of hominins closer to their common ancestor with chimpanzees, the Ardipithecus fossils strongly suggest, say the authors, that the common ancestor was substantially different in a number of ways from modern chimpanzees. Modern chimpanzees, and by implication gorillas as well, may have undergone substantial evolutionary changes of their own since their common ancestors with hominins. Although humans are certainly apes in the broad sense, the authors argue that modern apes do not provide a good analog for early human ancestors.
Ardipithecus appears to represent early hominins before they made the radical shift in adaptation from forest to a more mixed forest and savannah environment. This change, first seen perhaps in Australopithecus, or more likely in later hominins including Homo erectus and the “robust” australopithecines, led to many subsequent developments: extinction for the robust australopithecines and advanced intelligence and technology in the genus Homo.
Some of the key characteristics of Ardipithecus are described below, followed by a discussion of what these characteristics may imply for our understanding of early hominin evolution. In considering the evidence, it should be remembered that, as with any significant scientific discovery, the dialectical process of review and critique by colleagues followed by further research, then additional review and critique, and so on, will continue.
Some interpretations presented now may be altered by additional discoveries and analysis in the future. The fossilized bone specimens that form the basis for the analysis of Ardipithecus are fragmented, in some cases highly so. A number of the skeletal elements required extensive reconstruction. While the specialists who conducted this work are highly qualified, the process involves some degree of judgment. The recovery of additional specimens will help support or modify the current reconstructions. This is the normal course of paleontological research.
There has been critical comment within the anthropological community regarding the length of time, 15 years, between the recovery of the Ardipithecus fossil specimens and their publication in Science, a period during which other researchers did not have access to the material. Other important hominin finds, such as the Nariokatome Homo erectus, were made more widely available to colleagues shortly after discovery, permitting more extensive collaboration in research. Nevertheless, the large research team involved in the analysis of the Ardipithecus fossils and the substantial resulting publications are a significant contribution to the field.
The findings
The authors of the papers published in Science view two principal sets of characters, having to do with locomotion and diet, as indicating that Ardipithecus differed significantly from modern apes.
While chimpanzees and gorillas move across the ground by “knuckle walking,” Ardipithecus appears to have walked upright. The available evidence indicates that the foramen magnum, the opening at the base of the skull that allows the spinal chord to connect to the brain, was located more directly underneath the skull, as in later hominins, rather than relatively more toward the rear, as in modern apes, whose bodies lean forward when they walk on the ground. However, Ardi’s arms are still somewhat long, just shorter than those of modern apes, suggesting that the arms were still important for locomotion in trees.
 Figure 3. Hypothesized cladistic diagram of hominoid evolution (Click image for larger view)
Figure 3. Hypothesized cladistic diagram of hominoid evolution (Click image for larger view)Chimpanzee [Pan] and gorilla hands have become highly specialized to support the substantial weights of their bodies during tree climbing and have also evolved for knuckle walking. Importantly, Ardi’s hands do not show specialization for knuckle walking, which involves flexing the fingers and resting weight on the middle bone (phalange) of each finger, excluding the thumb. The authors argue that hominin hands, rather than losing the locomotor specializations of other apes, instead retained the more primitive, flexible form of earlier primates, which is better adapted to grasping objects. This flexibility, as well as the relative lengthening of the thumb that appears later, gave hominins the ability to manipulate objects in a variety of ways, using both a power grip and a precision grip. This, in turn, permitted the complex and precise use of tools.
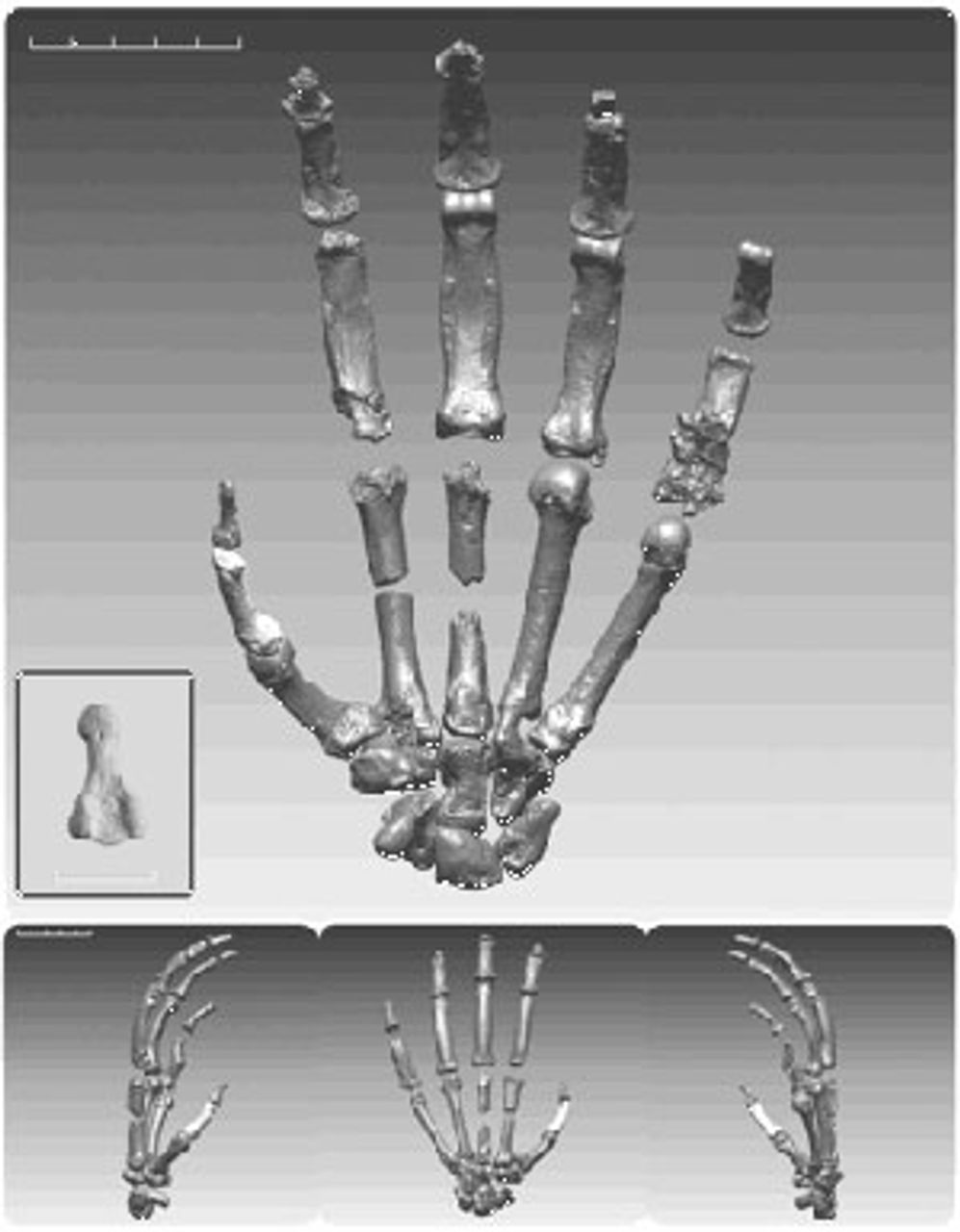 Figure 4. Reconstructed hand of an Ardipithecus
Figure 4. Reconstructed hand of an Ardipithecus specimen
The Ardipithecus pelvis is shorter and broader than is the case in modern apes. It represents a substantial advance toward the form seen in Australopithecus, a definite biped. If this interpretation is correct, then the ancestors of Ardi must have already been bipedal for a significant amount of time. This degree of evolutionary development would seem to imply that the split with the chimpanzee lineage may have occurred earlier than is currently thought. However, not all researchers agree on just how thoroughly Ardi was adapted for bipedalism.
Modern African apes have a foot highly adapted for tree climbing. It is much more specialized than that of Ardi and, argue the authors, more specialized than the foot of the last common ancestor. Ardi actually has a rigid foot structure reminiscent of monkeys, which the authors suggest was common to all ape ancestors, including those of both the modern African apes and hominins. This more rigid foot structure, originally adapted for leaping between branches, contend the authors, later proved useful to hominins since it improves the lever action of their feet while walking on the ground.
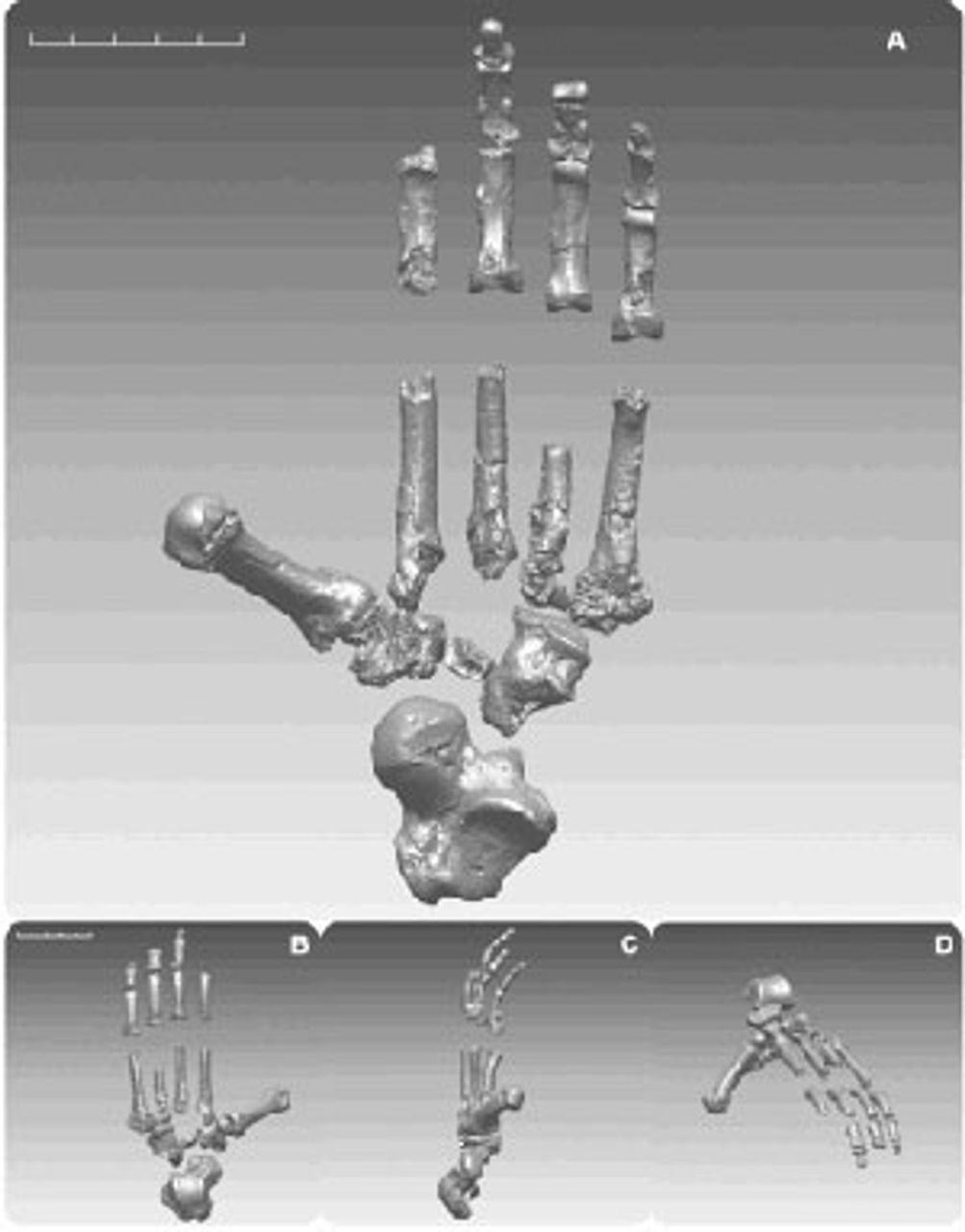 Figure 5. Reconstructed foot of an Ardipithecus
Figure 5. Reconstructed foot of an Ardipithecus specimen
If this is true, more-rigid feet were a “pre-adaptation” for the move by hominins from the forest to the savannah. However, Ardi’s big toes were spread significantly away from the line of the other toes, a feature that was likely useful for climbing trees, but less effective for walking on the ground. This and the retention of relatively long arms suggest that Ardipithecus spent time both in the trees and on the ground.
While Ardipithecus walked upright as we do, it had a cranial capacity (i.e., brain size) far below our own—of only about 300-350 cubic centimeters, approximately the same as that of modern apes of similar size and smaller than that of Australopithecus. It is possible that some reorganization of the brain may have taken place, perhaps beginning to enhance mental abilities beyond those of modern chimpanzees.
However, given the substantial increase in brain size among later hominins, associated with the development of culture and technology, it is likely that Ardipithecus had made no or only very limited advances in intelligence. Bipedalism and the consequent freeing of the hands had begun to evolve prior to the enlargement of the brain and the correlative expansion of intelligence. This “hands first” theory of human evolution was, for a long time, a minority view among anthropologists. One of its early supporters was Frederick Engels, who proposed it more than 130 years ago in his work The Part Played by Labor in the Transition from Ape to Man.
The other notable adaptation by Ardipithecus has to do with diet. These hominins do not have the large, thickly enameled post-canine teeth adapted for heavy chewing that are seen in Australopithecus. This suggests that the Ardipithecus had a varied diet that included a good proportion of softer foods such as ripe fruit, as opposed to Australopithecus, which was adapted to a diet containing tougher foods such as roots and seeds. This contrast likely reflects the difference between a creature that lived in a wooded environment and one that lived in a more mixed landscape.
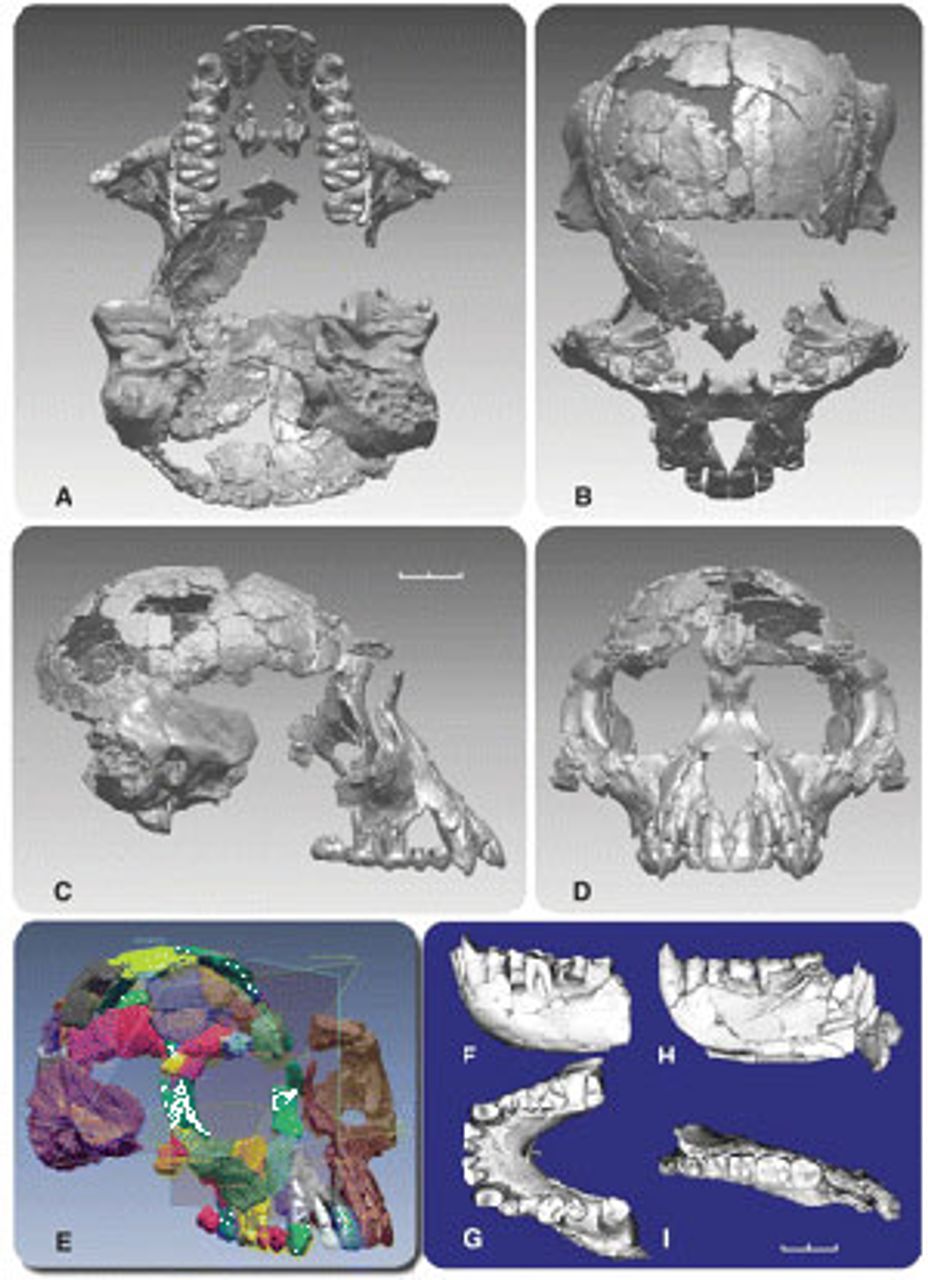 Figure 6. Reconstructed skull and teeth of an
Figure 6. Reconstructed skull and teeth of an Ardipiethcus specimen
Ardipithecus’s lower face (i.e., the mandible and maxilla, or “muzzle”) does not project as far forward as that of chimpanzees, but more than in Australopithecus. Chimps are primarily ripe-fruit eaters. The position of Ardipithecus’s tooth array more directly under the skull would result in more powerful chewing, since the muscles attached to the cranium would have a straighter up and down orientation than is the case for animals, such as chimps, with an extended muzzle.
Therefore, the Ardipithecus dentition and chewing apparatus may be seen as intermediate between that of chimpanzees, who eat relatively soft foods, and Australopithecus, which ate harder foods. This intermediate condition may reflect an equally intermediate diet that would have provided the flexibility to gradually shift to the savannah environment.
Ardipithecus’s canine teeth are significantly reduced compared to those of chimpanzees and gorillas. Reduction of canine size is a trend that was continued in later hominins. Two, not necessarily mutually exclusive explanations may account for this change. First, as foods that require grinding with the molars, such as seeds, became a larger part of the diet, large canines became an impediment to the necessary side-to-side motion.
The second possible factor in canine reduction may have to do with aggression within the social structure. Large canines are used by modern ape males in threat displays in order to gain social dominance. Males also tend to be significantly larger than females, also thought to be a product of social competition. The available evidence seems to indicate that there was little sexual dimorphism (i.e., difference between males and females) with respect to body size in Ardipithecus. The researchers suggest that the reduction in canine size may be related to a reduction in aggression between males and possibly marks the beginning of pair bonding between males and females. This proposal, by Owen Lovejoy of Kent State University, is one of the most controversial made in the series of articles in Science. It has been disputed by other researchers, notably David Pilbeam of Harvard, as being unsupported by the available data.
Interpretation
African apes and hominins both adapted to terrestrial locomotion from their primarily arboreal, monkey-like, common ancestors, but the methods adopted to solve the problem of moving effectively on the ground differed. Modern great apes, both gorillas and chimpanzees, use the “knuckle walking” mode while on the ground. These apes have arms longer than their legs. Therefore, when they walk on the ground, they are quadrupedal (using the ends of all four limbs as “feet”), but their bodies are held at a roughly 45-degree angle to the ground, as opposed to most quadrupedal animals, whose bodies are horizontal (i.e., parallel to the ground), on the one hand, and to hominins, who are bipedal, with their bodies held vertically, or perpendicular to the ground, on the other.
Hominins, by contrast, developed bipedalism as their way of moving on the ground. The reasons for this divergence (i.e., between knuckle walking and bipedalism), will require much more fossil evidence before they can be addressed effectively. The Ardipithecus fossils—and several even older but less extensive fossil finds, currently classified in different genera, Orrorin and Sahelanthropus—indicate that the evolution of bipedality must have begun very early after the split between hominins and the great apes. Indeed, bipedality is most likely one of the initial, defining characteristics of hominins.
Data collected along with the Ardipithecus fossils indicates that the environment in which these early hominins lived was characterized by woodland or forest patches, but not tropical rainforest. Therefore, it would not always have been possible for them to move from one tree to another without descending to the ground. It has been hypothesized that early hominins diverged from the great apes because they adapted to the increasingly open environment by developing bipedality in order to cross the growing distances from one tree to the other. The great apes in their turn may have maintained an adaptation to forested areas and, consequently, their habitats shrank along with the forests.
The new Ardipithecus data contributes to a growing consensus that, while this hypothesis is likely true in a broad sense, the actual evolutionary process was more complex and drawn out. The evidence for this more detailed interpretation comes in two main categories: locomotion and diet.
The Ardipithecus fossils indicate that the hominin method of locomotion began to evolve very early after the split with great apes and was already well on its way by 4.4 mya. The evidence of bipedality in Ardipithecus is most clearly seen in modifications of the pelvis (low and wide as opposed to long and narrow, as in apes) and associated musculature, based on the positions of muscle attachments to the bones and on the position of the foramen magnum underneath the skull, rather than to the rear. These trends would become even more developed in Australopithecus.
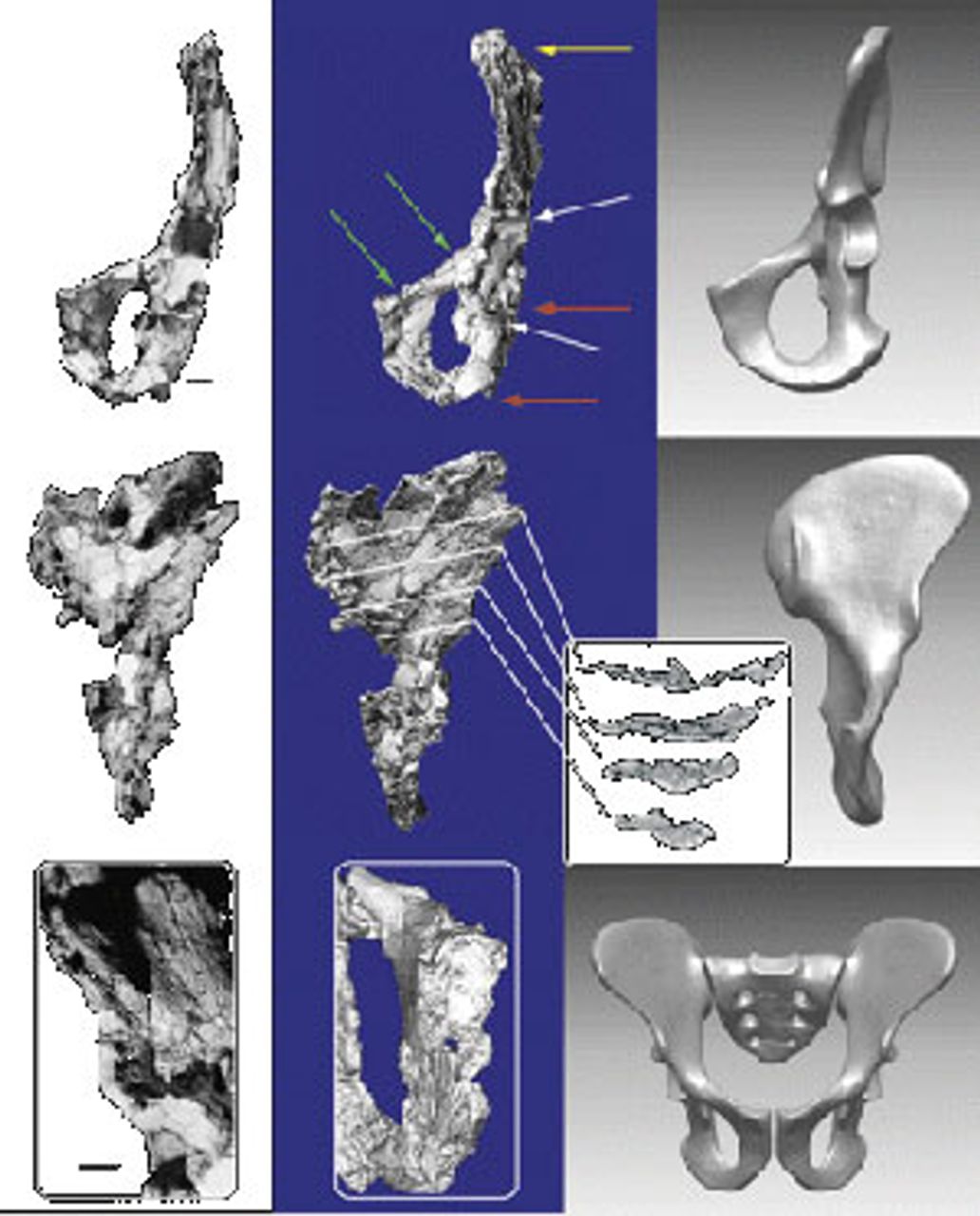 Figure 7. Pelvis of an Ardipiethcus specimen
Figure 7. Pelvis of an Ardipiethcus specimenBecause remains do not easily fossilize in heavily wooded or tropical regions, we do not have a powerful fossil lineage for modern apes and cannot be sure exactly how ape ancestors, including our own ancestors, moved in trees. However, it is safe to say that the common ancestor was primarily arboreal and, therefore, bipedality is a derived (i.e., newly evolved) adaptation not characteristic of the root lineage.
The other principal characteristics that differentiate hominins from great apes have to do with eating, specifically attributes of the teeth, maxilla and mandible (upper and lower jaws, respectively), and the associated musculature. The authors argue that hominin omnivory, allowing them to exploit a wide variety of food resources both in the trees and on the ground, is retained from pre-ape (i.e., monkey-like) and early ape ancestors. Although great apes, especially chimpanzees, have some flexibility in their diets, they became primarily specialized to eating forest foods, predominantly, though not exclusively, fruits and other forest vegetation.
This divergence in diet must have started with the beginning of the climatic drying-out process described above. Thus, as the forests slowly retreated and increasingly open environments developed along their margins, the relative diversity of the ancestral diet permitted the flexibility for local populations to tweak their food intake to adapt to local conditions.
Some populations of ancestral apes began placing relatively more emphasis on forest-based food sources and other populations leaned a little toward the more terrestrial and open ground sources. At first, this was simply “variations on a common theme.” Gradually, however, as environmental differentiation increased, both ecologically and geographically, along a spectrum from dense forest to open grassland, these various ancestral ape populations became more widely separated.
The shifts in diet were conservative in the sense that they were not radical adoptions of entirely new diets, but merely changes in relative proportions of the different constituents, while attempting to retain a diversified menu. For hominins, the unexpected combination of characteristics found in Ardipithecus, including bipedality on the ground and attributes useful for tree-climbing, represents the intermediate locomotor morphology needed to obtain this mixed diet. The intermediate position of Ardipithecus is supported by chemical evidence. Ardipithecus remains are found to contain ratios of heavy to light carbon that resemble woodland animals like chimpanzees, but are slightly closer to australopithecine ratios common among grassland animals.
The growing spatial separation between the locations of food resources and locations for sleeping and protection from predators may have favored bipedal locomotion. For early hominins who were slowly increasing their consumption of open-ground foods, such as grass seeds, it would have been an advantage to be able to collect and then carry foods back to the safety of the trees, rather than having to sit in the open while consuming them. Bipedality frees the hands for carrying, and observations of free-ranging chimpanzees indicate that their limited amount of bipedal locomotion is often associated with carrying food or objects.
The characteristics of Ardipithecus dentition and configuration of their jaws suggest an adaptation to grinding of harder foods, such as seeds, as opposed to the slicing and chopping action needed for eating fruits and vegetation, as seen in the great apes. By the time of Lucy (Australopithecus afarensis), 3.2 mya, the changes both in dentition/chewing apparatus and in locomotion seen in their early stages in Ardipithecus had become substantially more well developed. The qualitative transition to an open-environment-dwelling ape (i.e., a hominin) had been effected.
Whether Ardipithecus was the direct ancestor of Australopithecus or an evolutionary side branch can only be determined by additional fossil finds. However this question is ultimately resolved, the Ardipithecus specimens raise the possibility that the previous conception that hominins evolved from a chimpanzee-like ancestor may not be correct. The evolution of hominins and great apes may have followed paths different from what has hitherto been hypothesized. Now that the Ardipithecus data has been published, the debate between those who envision a chimpanzee-like hominin origin and those who do not will undoubtedly progress, providing a richer understanding of human evolution.
Related pamphlet: The Part Played by Labour in the Transition from Ape to Man by Frederick Engels